Pagina Inhoud:
Inleiding:
De motor-eindplaat is de verbinding tussen de motorische zenuw en de skeletspier cel. Zijn functie is om een elektrisch signaal (= actiepotentiaal) van de zenuwcel naar een spiercel over te brengen. Dit is de prikkel waardoor de skeletspier zal samentrekken (=contraheren). Een andere naam voor de motorische eindplaat is ‘neuromusculaire junctie’ (‘neuro’ = zenuw en ‘musculair’ = spier).
De motor-eindplaat is de verbinding tussen de motorische zenuw en de skeletspier cel. Zijn functie is om een elektrisch signaal (= actiepotentiaal) van de zenuwcel naar een spiercel over te brengen. Dit is de prikkel waardoor de skeletspier zal samentrekken (=contraheren). Een andere naam voor de motorische eindplaat is ‘neuromusculaire junctie’ (‘neuro’ = zenuw en ‘musculair’ = spier).
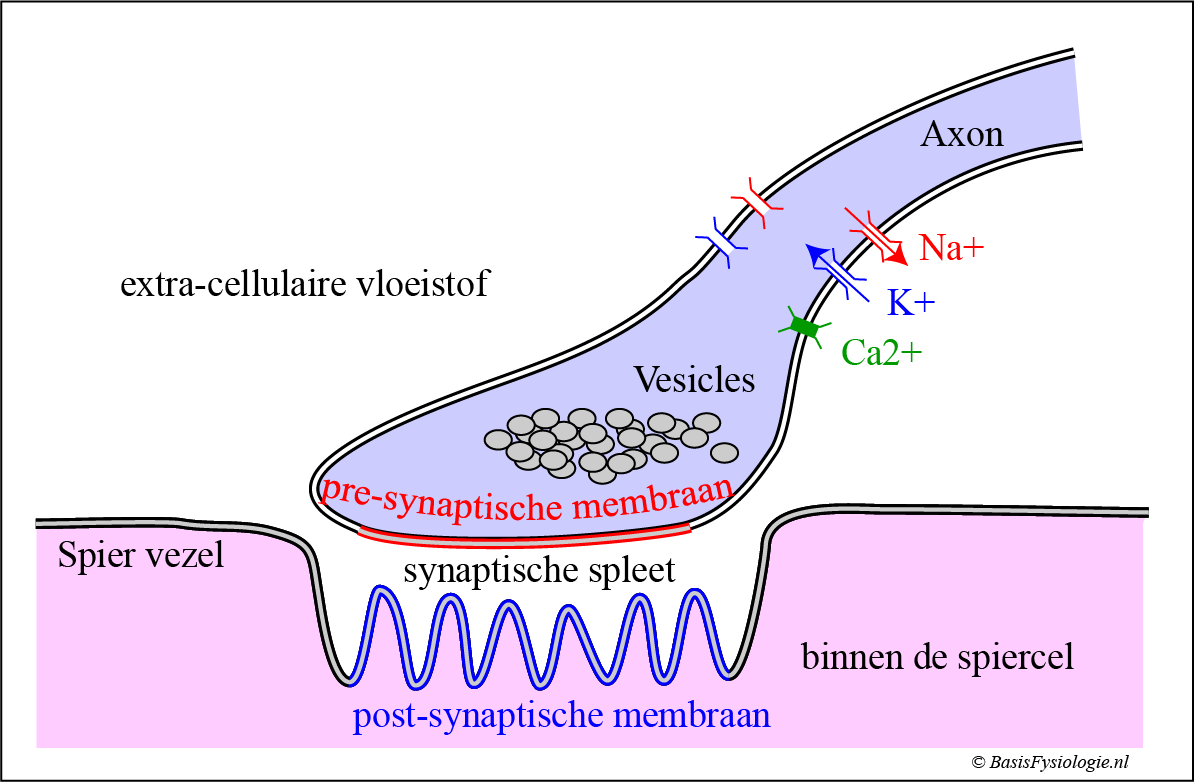
A. Structurele componenten van de motorische eindplaat:
- Een presynaptisch membraan in het distale deel van het axon.
- Dit presynaptische membraan bevat Na+, K+ en Ca2+ ion kanalen.
- Blaasjes in de presynaptische cel die de (neuro) transmitter bevat.
- In de motor-eindplaat is de neurotransmitter altijd acetylcholine (ACh)
- Een post-synaptisch membraan aan de spiercel. Deze zijn meestal gevouwen (raad eens waarom? Zie paneel H)
- Receptor operated kanalen (= ROC) die zich in het post synaptische membraan bevinden.
1.
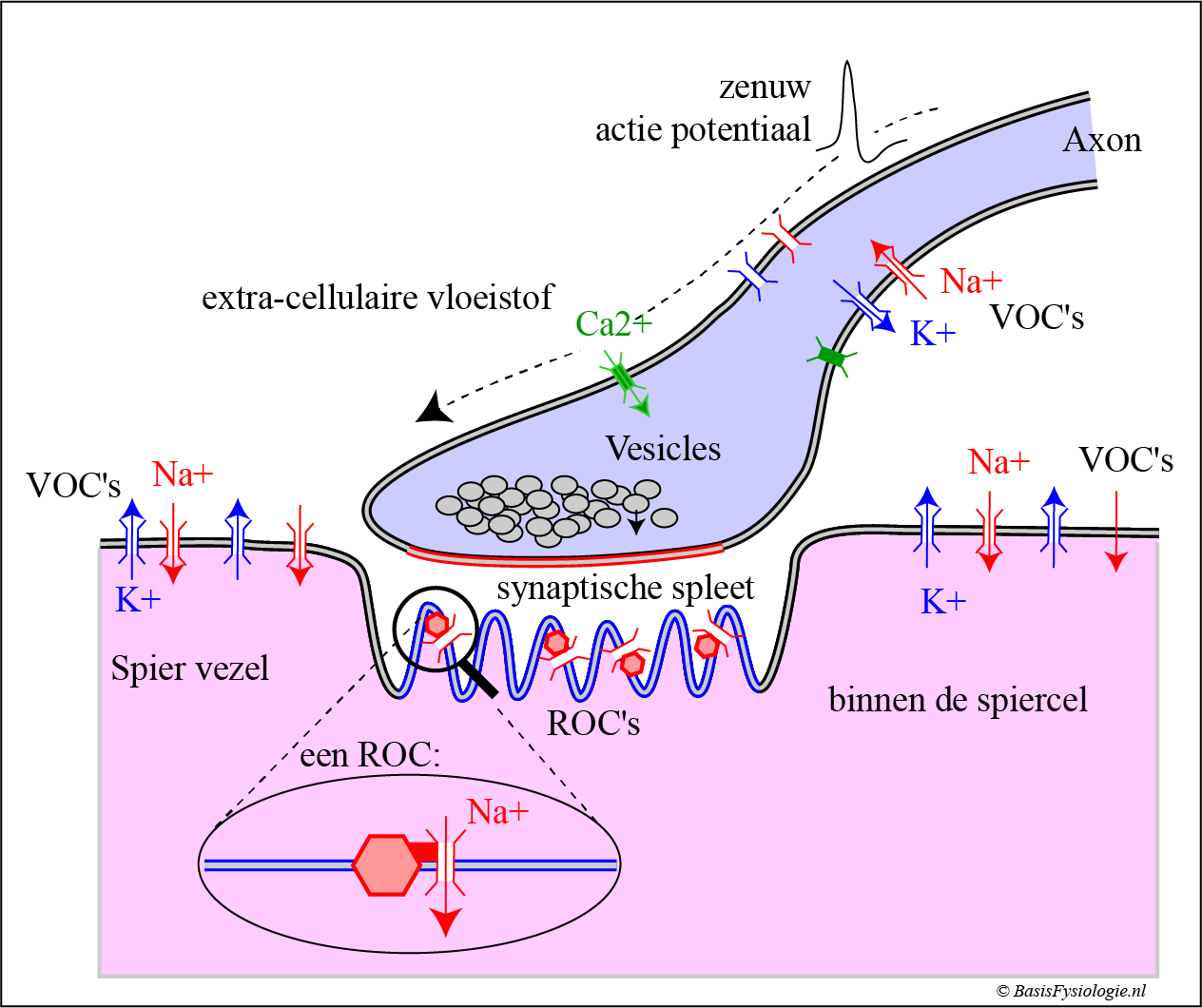
Een zenuw actiepotentieel geleidt, vanuit de zenuwcel, via de axon naar het presynaptische membraan.
2.
Deze actiepotentiaal, in het laatste deel van het axon, opent de Ca2+-kanalen.
3.
Vanwege de concentratiegradiënt (meer calcium buiten en minder binnen), zullen calciumionen de cel in stromen.
Vanwege de concentratiegradiënt (meer calcium buiten en minder binnen), zullen calciumionen de cel in stromen.
4.
Dit intracellulaire calcium zal één van de blaasjes (=vesikel) ertoe aanzetten zich naar het presynaptische membraan te verplaatsen.
5.
Eenmaal bij het presynaptische membraan aangekomen zal het blaasje versmelten met het membraan en de inhoud (acetylcholine) uitstorten in de synaptische spleet. Dit proces wordt exocytose genoemd.
6.
De transmitter moleculen (AcetylCholine = ACh) diffunderen in de synaptische spleet en sommige moleculen bereiken het post-synaptische membraan.
7.
Deze acetylcholine moleculen zullen dan koppelen aan de specifieke receptoren (ACh-receptoren) die zich in het post-synaptische membraan bevinden.
8.
De ACh-receptoren zijn gekoppeld aan ionenkanalen (receptor operated channels = ROC).
9.
De koppeling van acetylcholine aan de ACh-receptor zal deze specifieke kanaal openen.
10.
Naarmate er meer en meer transmitters zich aan de receptoren hechten, zullen steeds meer kanalen worden geopend.
11.
Het openen van deze kanalen veroorzaakt een stroom van Na+ -ionen in de post-synaptische cel (= influx).
12.
Deze positieve ionen zullen lokaal een depolarisatie van dat membraan veroorzaken
13.
Dit potentieel wordt een generator potentiaal genoemd.
14.
Wanneer de generatorpotentiaal de drempel bereikt, wordt een actiepotentiaal gegenereerd.
Wanneer de generatorpotentiaal de drempel bereikt, wordt een actiepotentiaal gegenereerd.
15.
De actiepotentiaal, eenmaal geïnitieerd, geleidt zich dan langs het spiermembraan rondom de spiercel en in de transversale tubuli.
16.
Dit zal het proces van contractie in de sarcomeren starten (zie volgende pagina).
1.
In de synaptische spleet, tussen de pre- en post-synaptische membranen, bevindt zich een enzym, acetylcholinesterase (= AChE).
2.
Deze acetylcholinesterase breekt het acetylcholine af.
3.
Deze afbraak is een noodzakelijke stap om te voorkomen dat de ACh continu aan de ACh-receptoren gekoppeld blijft.
4.
Zonder dit enzym zou het post-synaptische membraan constant worden gedepolariseerd, waardoor nieuwe actiepotentialen niet meer zou kunnen worden geïnduceerd.
1.
Het is belangrijk te beseffen dat het afbreken van de transmitter plaatsvindt in de synaptische spleet. Dit is eigenlijk in de extracellulaire ruimte (= buiten de cel).
2.
Dit maakt het gemakkelijk om dit mechanisme te beïnvloeden door een gif of medicijnen.
3.
Curare is bijvoorbeeld een bekend gif dat concurreert met acetylcholine (ACh) om de door de receptor bediende kanalen te bezetten.
Curare is bijvoorbeeld een bekend gif dat concurreert met acetylcholine (ACh) om de door de receptor bediende kanalen te bezetten.
4.
Maar in tegenstelling tot ACh opent curare de kanalen niet en veroorzaakt daarom géén generatorpotentiaal.
5.
Aangezien curare geen generatorpotentiaal induceert, induceert het dus geen actiepotentialen en daarom zullen er geen contracties zijn. In feite induceert curare verlamming !
6.
Curare werd door de Zuid-Amerikaanse Indianen gebruikt om hun prooien te doden (en ook mensen!). De vraag is nu waarom curare een zoogdier (zoals mensen) doodt?
Curare werd door de Zuid-Amerikaanse Indianen gebruikt om hun prooien te doden (en ook mensen!). De vraag is nu waarom curare een zoogdier (zoals mensen) doodt?
7.
Op dit punt aangekomen, zullen studenten vaak zeggen dat curare ook het hart blokkeert.
8.
Maar het hart is géén skeletspier, heeft geen motorische eindplaten, dus curare kan deze niet-bestaande ACh-receptoren niet blokkeren.
Maar het hart is géén skeletspier, heeft geen motorische eindplaten, dus curare kan deze niet-bestaande ACh-receptoren niet blokkeren.
9.
Maar de ademhaling in het lichaam wordt uitgevoerd door de skeletspieren van de borstkast. En we hebben het hier over de skeletspieren.
10.
Dus curare verlamt ook de spieren van de ademhaling en daarom wordt de ongelukkige prooi gedood door verstikking!!
Dus curare verlamt ook de spieren van de ademhaling en daarom wordt de ongelukkige prooi gedood door verstikking!!
11.
Tegenwoordig heeft de farmaceutische industrie curare-mimetica ontwikkeld (mimetica = werkt als curare).
12.
Met deze medicijnen kan men tijdelijk alle skeletspieractiviteit blokkeren. Dit is bijvoorbeeld handig tijdens een operatie.
13.
Maar natuurlijk moet men er ook voor zorgen dat de ademhaling niet wordt gestopt. Omdat de ademhalingsspieren verlamd zijn, wordt de ademhaling tijdens een operatie overgenomen door een mechanische ventilator.
14.
En, misschien nog belangrijker, is om de patiënt bewusteloos te maken.
15.
Omdat, als de patiënt tijdens de operatie niet bewusteloos is, de patiënt in feite verlamd is maar niet kan schreeuwen, bewegen of praten!
16.
Dit is natuurlijk een nachtmerrie!
Dit is natuurlijk een nachtmerrie!

1.
Misschien is het goed om hier nog even in te gaan op het verschil tussen ROC’s en VOC’s. In een vorige pagina heb je al kennis gemaakt met ROC’s; werkzaam op het post-synaptische membraan van de chemische synaps (A.3.7. De Chemische Synaps).
2.
In feite werken de ROC’s op de spiercel op dezelfde manier. Maar belangrijk is ok het verschil tussen ROC’s en VOC’s (die o.a. voor de actiepotentiaal geleiding zorgen).
3.
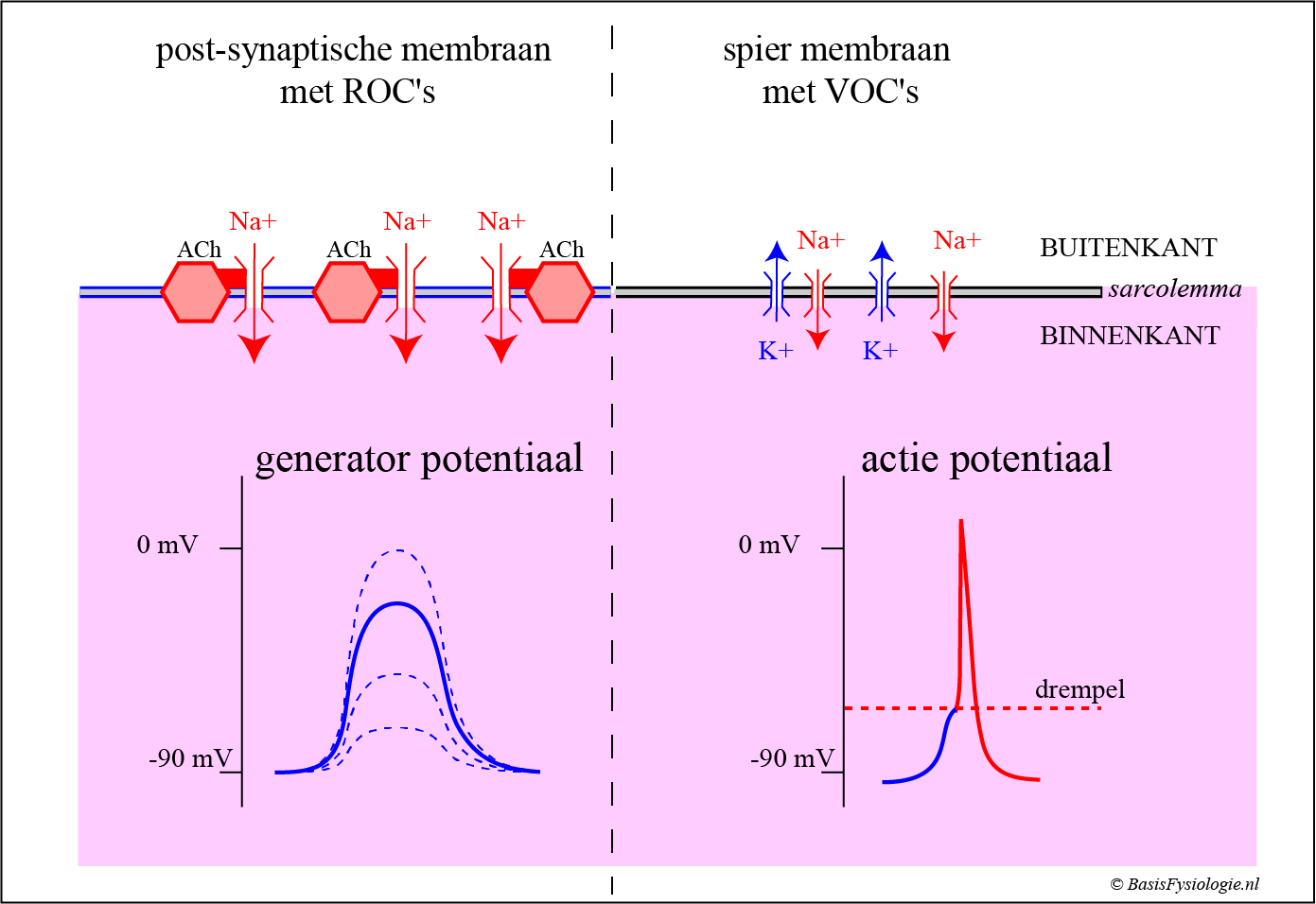
Receptor-operated channels worden ‘operated’ wanneer de transmitter (=ACh) gekoppeld wordt aan de receptor.
4.
Voltage-operated channels worden ‘operated’ door de transmembraan potentiaal (dit is het stroomverschil over het membraan; in rust negatief binnen en positief buiten).
Voltage-operated channels worden ‘operated’ door de transmembraan potentiaal (dit is het stroomverschil over het membraan; in rust negatief binnen en positief buiten).
1.
Een generatorpotentiaal geleid niet en is daarom een lokaal fenomeen (terwijl de actiepotentialen wel geleiden).
2.
Een generatorpotentiaal is gradueel; het is klein wanneer slechts enkele transmitters aan hun receptoren zijn gekoppeld en wordt groter wanneer meer transmitters zijn aangesloten (dus niet beperkt door de beroemde Alles-of-Niets wet!).
Een generatorpotentiaal is gradueel; het is klein wanneer slechts enkele transmitters aan hun receptoren zijn gekoppeld en wordt groter wanneer meer transmitters zijn aangesloten (dus niet beperkt door de beroemde Alles-of-Niets wet!).
3.
Dus, als de generatorpotentiaal de drempel niet bereikt, dan ontstaat er géén actiepotentiaal.
4.
De generatorpotentiaal heeft ook geen refractaire periode, terwijl een actiepotentiaal dat wel heeft.
1.
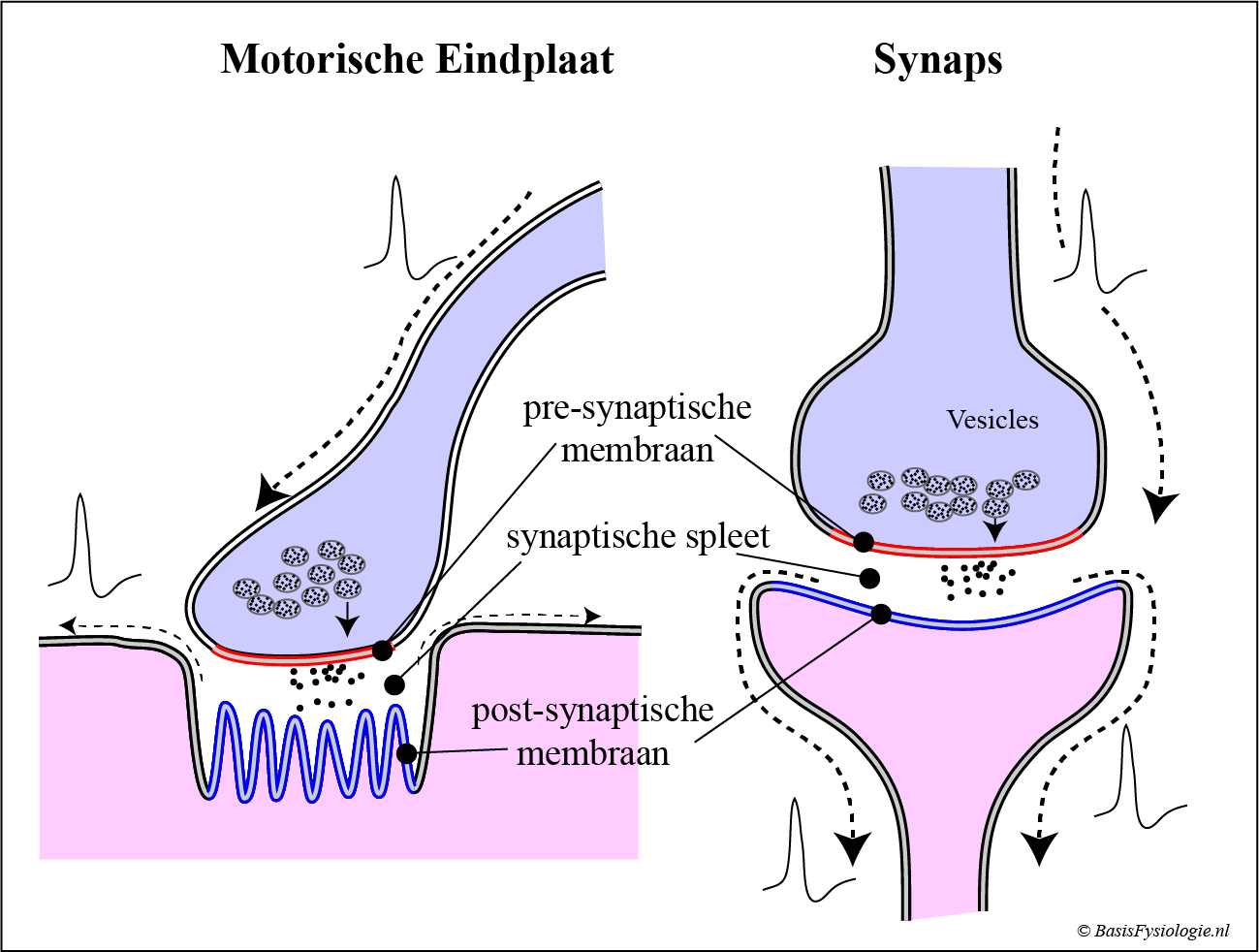
Op een vorige pagina zagen we de structuur en functie van een chemische synaps; de verbinding tussen twee zenuwcellen.
2.
Op deze pagina hebben we een ander soort chemische synaps geïntroduceerd, de motorische eindplaat, die een zenuwcel verbindt met een skeletspiercel.
Op deze pagina hebben we een ander soort chemische synaps geïntroduceerd, de motorische eindplaat, die een zenuwcel verbindt met een skeletspiercel.
3.
Er zijn een aantal belangrijke verschillen tussen deze twee structuren die we hier moeten bespreken.
4.
Het belangrijkste structurele verschil is dat het post-synaptische membraan in de motor-eindplaat sterk gevouwen is (waarom?), wat meestal niet het geval is in een chemische synaps.
5.
Cruciale verschillen tussen synaps en motor-eindplaat zijn:
- soort transmitters
- 1: 1
- Generatorpotentieel versus IPSP of EPSP
6.
In een motorische eindplaat is de transmitter altijd acetylcholine (= ACh). In de synaps kan dit een van de vele (neuro-) transmitter zijn (ACh, adrenaline, DOPA, enz.).
7.
De transmitters in de motor-eindplaat creëren altijd een generatorpotentiaal die het membraan depolariseert naar de drempel toe.
8.
In de chemische synaps worden, afhankelijk van het type transmitter, depolariserende (= EPSP) of hyper polariserende (= IPSP) stromen gegenereerd (Zie: ‘Chemische synaps’)
In de chemische synaps worden, afhankelijk van het type transmitter, depolariserende (= EPSP) of hyper polariserende (= IPSP) stromen gegenereerd (Zie: ‘Chemische synaps’)
9.
Ten slotte bereikt een generatorpotentiaal in de motorische eindplaat altijd de drempelwaarde (en initieert dan een actiepotentiaal in de spiercel). Daarom is de overdracht van een actiepotentiaal naar een skeletspiercel altijd succesvol. De verhouding is daarom 1: 1.
10.
Zoals besproken (‘Chemische synaps’), is dit in de synaps niet het geval. Daar is de verhouding van pre- tot post-synaptische actiepotentialen typisch 1:10 is.
1.
Door het membraan op die locatie te vouwen, wordt het oppervlak van dat deel van het membraan vergroot.
2.
Een groter oppervlak betekent dat er meer ROC’s op dit membraan kunnen worden geplaatst.
3.
Dus, wanneer ACh diffundeert van het presynaptische membraan in de synaps spleet, kan deze transmitter worden gekoppeld aan meer beschikbare ROC’s.
Dus, wanneer ACh diffundeert van het presynaptische membraan in de synaps spleet, kan deze transmitter worden gekoppeld aan meer beschikbare ROC’s.
4.
Dit zal op zijn beurt een grotere instroom van Na+ ionen creëren en daarom een groter generatorpotentieel induceren.
5.
Dit zorgt er dan ook voor dat er altijd een actiepotentiaal in de spiercel wordt gecreëerd.
6.
Kortom, de vouwen in de membraan maakt dit soort synaps veel gevoeliger en de geleiding betrouwbaarder!
